Ardipithecus Ramidus: The evolutionary consequences of SS
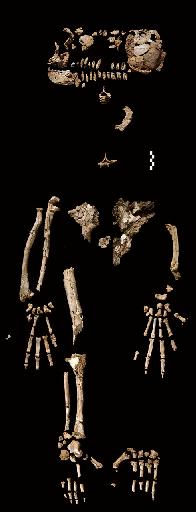
Following the long anticipated unveiling of the Ardipithecus Ramidus materials (Oct. 2009) [1], White et al. put-forth a new hypothesis to explain the acquisition of bipedality in hominids. Compared to fossil and modern apes, males of the Ardipithecus Ramidus species were shown to have a "significantly reduced size in their canine teeth". Under this hypothesis, two associated traits - reduced canine dentition and bipedality - carried by Ar. Ramidus males were selected for as "attractive reproductive traits" by the females of the species.
Paralleling Darwin in the Descent of Man [2], White et al. insist that - unlike in most ape species, where males compete with each other (using their canines as weapons) for access to females, Ar. Ramidus females chose less combative (by the indicator of reduced canine size) and more adequate bipedalist males as mates. Less combative males form more stable "monogamous, pair bonds", and more effective bipedalists transfer more resources to females and offspring (relative to more combative and less competent bipedalist males). [3]
White et al. raise the shift in a sexual selection mechanism from (apelike) "male-male competition" to (hominid-like) "female choice" to help unravel a persisting riddle: Why did hominid species diversify rapidly in Africa around 4 million years ago? Some of the descendants of this "hominid population explosion" acquired large brains, tool-making ability and articulate speech. Eventually, they spread out from Africa to colonize the entire planet.
Human females in male-female "pair bonds" (can) produce more offspring than polyandrous ape females - who are compelled to live in dominant male harems, when showing signs of ovulation. For example, chimpanzee females are fertile for a short period of time during the year (7-12 days), and they give birth to only one infant every 3 - 5 years. Human females are regularly fertile, and they (can) produce significantly more offspring than modern apes. It's postulated that - by hominid females offering males sex on a regular basis for food (and other resources), the hominid lineage experienced a population explosion. [4]
Over time, the associated traits of reduced canine dentition (as a reproductive advantage) and bipedality (as both a reproductive and a survival advantage) became "fixed" in the hominid line - which gave rise to humans. Less combative and better walking males out-produced - in terms of offspring - more violent and less agilely walking males. Reduced male canine size is to be understood as one of Darwin's "male ornaments", i.e. an attractive reproductive trait. For example, 4.5 Myr ago, (relatively) small canine teeth, carried by Ardi males in a forest teeming with predators, should not have conveyed an obvious survival advantage on them, but this trait - through Sexual Selection - conveyed what appears to have been a reproductive advantage.
One conspicuous flaw in the "choosey female hypothesis" is that evidence for a shift in sexual selection from "male combat" in fossil apes to "female choice" in fossil hominids +4 Myrs ago cannot be found directly (to a high degree of certainty) in the fossil record. (Differentiating a survival trait from a reproductive trait from the fossilized remains of an organism is problematic.) It can, however, be inferred - indirectly - from the traits of bipedalism, reduced canine size, the capacity of the hominid line to produce more offspring than apes and the appearance of the abrupt "spinning off" of closely-related hominid species after 4 Myrs ago.
In insect, fish, bird, other mammal, etc. species, even minor shifts in female mate preference can (and have been convincingly shown) to have led to radical, long term evolutionary consequences - to wit: An increase (or decrease) in sexual dimorphism and in a few cases a proliferation of daughter species. The new hypothesis makes more evolutionary sense - than previous hypotheses, invoked to account-for the appearance of hominid bipedality. The construction of the "choosey female hypothesis" resembles how evolutionary biologists confront and explain evolutionary breakthroughs and trends - in populations of non-human organisms.
Contrast the "choosey female hypothesis" with two previous hypotheses, advanced to explain the trait of bipedality in hominids: (1) the "savanna" and (2) the "brain cooler" hypotheses. Popularized in the 1970s - after the discovery of Lucy (Australopithecus Afarenis) by Don Johanson, the "savanna hypothesis" runs that a survival advantage was conferred on those hominids having the ability to "peer over" tall grasses in evading predators or locating food. The "savanna hypothesis" began to breakdown in the early 1990s - when at the time - sketchy evidence emerged that hominids walked upright prior to the expansion of the African savanna (about 3.5 Myr). As a result, an older "brain cooler" hypothesis was dusted-off, revised and hauled-out to describe the achievement of bipedalism. [5, 6] Walking upright cools an increasing in size brain. A relatively enlarged brain is a massive heat generator. Bipedalism, under the "brain cooler" hypothesis, is said to have conferred a survival advantage on bipedalists - since less surface area of their bodies would have been exposed to the hot, African sun than on quadrupedalists, militating-against the possibility of fatal heat-strokes.
The individuals of Ar. Ramidus lived in the forest and walked upright, so the "savanna hypothesis" is more than problematic. It's refuted. Ardi's brain-size is estimated to have been smaller than the size of a chimpanzee's brain, so the "brain cooler hypothesis" fits neither Ardi's cranial features nor (probably) the ecological conditions under which bipedality arose. The "choosey female hypothesis" ties multiple traits, patterns and trends in hominid evolution together seamlessly, giving it a greater explanatory power than other competing hypotheses.
Contrast again the approach of White et al. with that of Louis Leakey. [7] In 1964 - at Olduvai Gorge, Leakey unearthed sets of tool-kits and specimens of two hominid species - one of which he believed ancestral to humans. He (and Raymond Dart) named this putative, ancestral species Homo Habilis. Habilines made tools, had largish brains (and by fragmentary evidence in 1964) walked up-right. (Some Habilis specimens have been reclassified as Homo Rudolfensis - a species from which humans did not descend). Leakey, also, pronounced that tool-making was "the hall-mark" of what it meant to be assigned to the Homo genus - because this trait is what makes our genus "special". Leakey's approach to looking at hominid species conflicts starkly with how evolutionary biologists look at other mammal, bird, insect, etc. species. To them, no genus requires an "ordinal trait" to prove it "special". What they study are anything-but-special organisms in nature and experiments.
Evolutionary biologists take Darwin at face-value. As an example: In experiments conducted on fruit flies, female fruit flies have been shown to prefer mating with males carrying high concentrations of a specific pheromone - rather than with males carrying lower concentrations. This "secondary sex trait" conveys signals of genetic quality to females. The males emitting more of a particular pheromone out-breed males who emit less of it. They and their descendants' genes can swamp the population and shift the genetic profile (first of the cline then the entire species), triggering long-term evolutionary trends. [8]
The hypothesis of White et al. takes Darwin more at face-value - than other bipedalist hypotheses. As I've suggested - due to the "choosey female hypothesis", finally paleo-anthropology has become "fully Darwinized".
"We should never have doubted Darwin in his appreciation that the ultimate source of our matchlessness among mammals would prove commonplace when knowledge became sufficiently advanced. Ar. ramidus now enhances that knowledge. Even our species-defining cooperative mutualism can now be seen to extend well beyond the deepest Pliocene." -- Owen Lovejoy from Reexamining Human Origins in Light of Ardipithecus ramidus
The emergences of reduced canine size and up-right walking - as reproductive traits in the hominid lineage - can be traced to Ar. Ramidus females, preferring to mate with Ar. Ramidus males - who "displayed" these "elaborate male traits". Ardi was in no way "special". He was forest-adapted, he had a large, opposable big toe, a puny brain and canine teeth-size intermediate in size between fossil apes and Lucy.
Will the "choosey female hypothesis" hold-up over time - as the explanation for the acquisition of bipedality in hominids? Every 10 to 15 years, the reigning scenario of hominid origins shifts, and the old scenario displaced. I predict that a paleo-anthropologist will "prove" that Ardi was not ancestral to Lucy - due to a new find or a re-interpretation of the Ardi materials, and the bold assertion that +4 Myr ago hominid males were "monogamous" and "pair-bonded" stretches credibility to the breaking point. Further, there is fragmentary evidence that the traits of reduced male canine size and bipedalism predated Ar. Ramidus by a million years or so.
Notes
(1) From Ardipithecus ramidus and the Paleobiology of Early Hominids:
"The adoption of bipedality and its temporal association with progressive canine reduction and loss of functional honing now constitute the principal defining characters of Hominidae." -- Tim D. White, Berhane Asfaw, Yonas Beyene, Yohannes Haile-Selassie, C. Owen Lovejoy, Gen Suwa and Giday WoldeGabriel
(2) The Descent of Man, and Selection in Relation to Sex, by Charles Darwin (1871) To be historically accurate, Darwin speculated that the mechanism of sexual selection in the human species operates by "male choice" - not "female choice". Darwin was a bit too reductive in applying his theory of sexual selection to humans. Darwin hypothesized that human females "display" the "ornaments" - such as large breasts, shapely legs and buttocks, full lips, etc. - which males select as attractive reproductive traits. But, his (more general) point remains well taken. There was a shift from "male combat" in ape species to "choice" (though female) in hominids.
(3) From Reexamining Human Origins in Light of Ardipithecus ramidus:
"The great apes are an isolated, uniquely specialized relict species surviving today only by their occupation of forest refugia." "Ar. ramidus now confirms that extant African ape-based models [of hominid evolution] are no longer appropriate." "[] at the base of the hominid clade, temporary pair bonds based on sex-for-food exchanges would have further encouraged copulation with provisioning males, rather than males that relied on dominance or aggressive displacement of competitors abetted by large and projecting canines. Research has confirmed the selective advantages of such exchanges []" -- C. Owen Lovejoy (2009)
(4) From Reexamining Human Origins in Light of Ardipithecus ramidus
"The perspective offered by Ardipithecus suggests that our special cognitive abilities derive from a unique earlier interplay of otherwise commonplace elements of locomotion, reproductive biology, neurophysiology, and social behavior." "We can no longer rely on homologies with African apes for accounts of our origins and must turn instead to general evolutionary theory." "A proposed adaptive suite for the emergence of Ardipithecus from the last common ancestor that we shared with chimpanzees accounts for these principal ape/human differences, as well as the marked demographic success and cognitive efflorescence of later Plio-Pleistocene hominids." -- Lovejoy
(5) The Fossil Trail, by Ian Tattersall (1995)
(6) Development of Bipedalism in Hominins, by Emma Wiley (2008)
(7) A new species of the Genus Homo from Olduvai Gorge, by Dr. L.B.S Leakey (1964)
(8) A Drosophila male pheromone affects female sexual receptivity, by Micheline Grillet, Laurence Dartevelle, and Jean-François Ferveur (2005)